Trilobites are a well-known fossil group of extinct marine arthropods that form the class Trilobita. Trilobites form one of the earliest known groups of arthropods. The first appearance of trilobites in the fossil record defines the base of the Atdabanian stage of the Early Cambrian period (521 million years ago), and they flourished throughout the lower Paleozoic era before beginning a drawn-out decline to extinction when, during the Devonian, all trilobite orders except Proetida died out. Trilobites finally disappeared in the mass extinction at the end of the Permian about 250 million years ago. The trilobites were among the most successful of all early animals, roaming the oceans for over 270 million years.
When trilobites first appeared in the fossil record they were already highly diverse and geographically dispersed. Because trilobites had wide diversity and an easily fossilized exoskeleton an extensive fossil record was left behind, with some 17,000 known species spanning Paleozoic time. The study of these fossils has facilitated important contributions to biostratigraphy, paleontology, evolutionary biology and plate tectonics. Trilobites are often placed within the arthropod subphylum Schizoramia within the superclass Arachnomorpha (equivalent to the Arachnata), although several alternative taxonomies are found in the literature.
Trilobites had many life styles; some moved over the sea-bed as predators, scavengers or filter feeders and some swam, feeding on plankton. Most life styles expected of modern marine arthropods are seen in trilobites, with the possible exception of parasitism (where there are still scientific debates). Some trilobites (particularly the family Olenidae) are even thought to have evolved a symbiotic relationship with sulfur-eating bacteria from which they derived food.
Fossil record

.and bigotinids (order Ptychopariida, superfamily Ellipsocephaloidea) dated to some 540 to 520 million years ago. Contenders for the earliest trilobites include Profallotaspis jakutensis (Siberia), Fritzaspis spp. (western USA), Hupetina antiqua (Morocco) and Serrania gordaensis (Spain). All trilobites are thought to have originated in present day Siberia, with subsequent distribution and radiation from this location.
Origins
Early trilobites show all the features of the trilobite group as a whole; there do not seem to be any transitional or ancestral forms showing or combining the features of trilobites with other groups (e.g. early arthropods).
Morphological similarities between trilobites and early arthropod-like creatures such as Spriggina, Parvancorina, and other “trilobitomorphs” of the Ediacaran period of the Precambrian are ambiguous enough to make detailed analysis of their ancestry far from compelling. Morphological similarities between early trilobites and other Cambrian arthropods (e.g. the Burgess Shale fauna and the Maotianshan shales fauna) make analysis of ancestral relationships difficult.
However, it is still reasonable to assume that the trilobites share a common ancestor with other arthropods before the Ediacaran-Cambrian boundary. Evidence suggests that significant diversification had already occurred before trilobites were preserved in the fossil record, easily allowing for the “sudden” appearance of diverse trilobite groups with complex derived characteristics (e.g. eyes).
Divergence and extinction
For such a long-lasting group of animals, it is no surprise that trilobite evolutionary history is marked by a number of extinction events where some groups perished while surviving groups diversified to fill ecological niches with comparable or novel adaptations. Generally, trilobites maintained high diversity levels throughout the Cambrian and Ordovician periods before entering a drawn-out decline in the Devonian culminating in final extinction of the last few survivors at the end of the Permian period.
Evolutionary trends
Principal evolutionary trends from primitive morphologies (e.g. eoredlichids) include the origin of new types of eyes, improvement of enrollment and articulation mechanisms, increased size of pygidium (micropygy to isopygy) and development of extreme spinosity in certain groups. Changes also included narrowing of the thorax and increasing or decreasing numbers of thoracic segments. Specific changes to the cephalon are also noted; variable glabella size and shape, position of eyes and facial sutures & hypostome specialization. Several morphologies appeared independently within different major taxa (e.g. eye reduction or miniaturization).
Effacement is also a common evolutionary trend. It is the loss of surface detail in the cephalon, pygidium, or the thoracic furrows. Notable examples of this were the orders Agnostida, Asaphida, and the suborder Illaenina of Corynexochida. It is believed that effacement is an indication of either a burrowing lifestyle or a pelagic one. Effacement poses a problem for taxonomists since the loss of details (particularly of the Glabella) can make the determination of phylogenetic relationships difficult.
Pre-Cambrian
Phylogenetic biogeographic analysis of Early Cambrian Olenellidae and Redlichidae suggests that a uniform trilobite fauna existed over Laurentia, Gondwana and Siberia before the tectonic breakup of the super-continent Pannotia between 600 million years ago and 550 million years ago.Tectonic breakup of Pannotia then allowed for the diversification and radiation expressed later in the Cambrian as the distinctive olenellid province (Laurentia, Siberia and Baltica) and the separate Redlichid province (Australia, Antarctica and China). Breakup of Pannotia significantly predates the first appearance of trilobites in the fossil record, supporting a long and cryptic development of trilobites extending perhaps as far back as 700 million years ago or possibly further.
Cambrian
Very shortly after trilobite fossils appeared in the lower Cambrian, they rapidly diversified into the major orders that typified the Cambrian—Redlichiida, Ptychopariida, Agnostida and Corynexochida. The first major crisis in the trilobite fossil record occurred in the Middle Cambrian; surviving orders developed isopygus or macropygius bodies and developed thicker cuticles, allowing better defense against predators (see Thorax above). The end Cambrian mass extinction event marked a major change in trilobite fauna; almost all Redlichiida (including the Olenelloidea) and most Late Cambrian stocks went extinct. A continuing decrease in Laurentian continental shelf area is recorded at the same time as the extinctions, suggesting major environmental upheaval.
Ordovician

echinoderms, and graptolites with many groups appearing in the fossil record for the first time. Although intra-species trilobite diversity seems to have peaked during the Cambrian, trilobites were still active participants in the Ordovician radiation event with a new fauna taking over from the old Cambrian one.
The Ordovician marks the last great diversification period amongst the trilobites, very few entirely new patterns of organisation arose post-Ordovician; later evolution in trilobites was largely a matter of variations upon the Ordovician themes. By the Ordovician mass extinction vigorous trilobite radiation has stopped and gradual decline beckons.
Silurian and Devonian

Most Early Silurian families constitute a subgroup of the Late Ordovocian fauna. Few, if any, of the dominant Early Ordovician fauna survived to the end of the Ordovician, yet 74% of the dominant Late Ordovician trilobite fauna survived the Ordovician. Late Ordovician survivors account for all post-Ordovician trilobite groups except the Harpetida.
Carboniferous and Permian
The Proetida survived for millions of years, continued through the Carboniferous period and lasted until the end of the Permian (when the vast majority of species on Earth were wiped out). It is unknown why order Proetida alone survived the Devonian. The Proetida maintained relatively diverse faunas in deep water and shallow water, shelf environments throughout the Carboniferous. For many millions of years the Proetida existed untroubled in their ecological niche. An analogy would be today’s crinoids, which mostly exist as deep water species; in the Paleozoic era, vast ‘forests’ of crinoids lived in shallow near-shore environments.
Final extinction
Fossil distribution

Trilobites appear to have been exclusively marine organisms, since the fossilized remains of trilobites are always found in rocks containing fossils of other salt-water animals such as brachiopods, crinoids, and corals. Within the marine paleoenvironment, trilobites were found in a broad range from extremely shallow water to very deep water. Trilobites, like brachiopods, crinoids, and corals, are found on all modern continents, and occupied every ancient ocean from which Paleozoic fossils have been collected. The remnants of trilobites can range from the preserved body to pieces of the exoskeleton, which it sheds in the process known as ecdysis. In addition, the tracks left behind by trilobites living on the sea floor are often preserved as trace fossils.
There are three main forms of trace fossils associated with trilobites: Rusophycus; Cruziana & Diplichnites – such trace fossils represent the preserved life activity of trilobites active upon the sea floor. Rusophycus, the resting trace, are trilobite excavations involving little or no forward movement and ethological interpretations suggest resting, protection and hunting.Cruziana, the feeding trace, are furrows through the sediment, which are believed to represent the movement of trilobites while deposit feeding. Many of the Diplichnites fossils are believed to be traces made by trilobites walking on the sediment surface. However, care must be taken as similar trace fossils are recorded in freshwater and post Paleozoic deposits, representing non-trilobite origins.
Trilobite fossils are found worldwide, with many thousands of known species. Because they appeared quickly in geological time, and moulted like other arthropods, trilobites serve as excellent index fossils, enabling geologists to date the age of the rocks in which they are found. They were among the first fossils to attract widespread attention, and new species are being discovered every year.
A famous location for trilobite fossils in the United Kingdom is Wren’s Nest, Dudley in the West Midlands, where Calymene blumenbachi is found in the Silurian Wenlock Group. This trilobite is featured on the town’s coat of arms and was named the Dudley Bug or Dudley Locust by quarrymen who once worked the now abandoned limestone quarries. Llandrindod Wells, Powys, Wales, is another famous trilobite location. The well-known Elrathia kingi trilobite is found in abundance in the Cambrian age Wheeler Shale of Utah.
Spectacularly preserved trilobite fossils, often showing soft body parts (legs, gills, antennae, etc.) have been found in British Columbia, Canada (the Cambrian Burgess Shale and similar localities); New York State, U.S.A. (Ordovician Walcott-Rust quarry, near Russia, and Beecher’s Trilobite Bed, near Rome); China (Lower Cambrian Maotianshan Shales near Chengjiang); Germany (the Devonian Hunsrück Slates near Bundenbach) and, much more rarely, in trilobite-bearing strata in Utah (Wheeler Shale and other formations), Ontario, and Manuels River, Newfoundland and Labrador.The French palaeontologist Joachim Barrande (1799–1883) carried out his landmark study of trilobites in the Cambrian, Ordovician and Silurian of Bohemia, publishing the first volume of Système silurien du centre de la Bohême in 1852.
Morphology
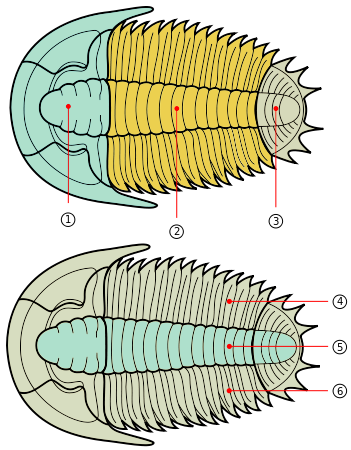
When trilobites are found, only the exoskeleton is preserved (often in an incomplete state) in all but a handful of locations. A few locations (Lagerstätten) preserve identifiable soft body parts (legs, gills, musculature & digestive tract) and enigmatic traces of other structures (e.g. fine details of eye structure) as well as the exoskeleton.
Trilobites range in length from 1 millimetre (0.04 in) to 72 centimetres (28 in), with a typical size range of 3–10 cm (1.2–3.9 in). The world’s largest trilobite, Isotelus rex, was found in 1998 by Canadian scientists in Ordovician rocks on the shores of Hudson Bay.
The exoskeleton is composed of calcite and calcium phosphate minerals in a protein lattice of chitin that covers the upper surface (dorsal) of the trilobite and curled round the lower edge to produce a small fringe called the “doublure”. Three distinctive tagmata (sections) are present: cephalon (head); thorax (body) and pygidium (tail).
Terminology
As might be expected for a group of animals comprising c. 5,000 genera, the morphology and description of trilobites can be complex. However, despite morphological complexity and an unclear position within higher classifications, there are a number of characteristics that distinguish the trilobites from other arthropods: a generally sub-elliptical, dorsal, chitinous exoskeleton divided longitudinally into three distinct lobes (from which the group gets its name); having a distinct, relatively large head shield (cephalon) articulating axially with a thorax comprising articulated transverse segments, the hindmost of which are almost invariably fused to form a tail shield (pygidium). When describing differences between trilobite taxa, the presence, size, and shape of the cephalic features are often mentioned.
During moulting, the exoskeleton generally split between the head and thorax, which is why so many trilobite fossils are missing one or the other. In most groups facial sutures on the cephalon helped facilitate moulting. Similar to lobsters and crabs, trilobites would have physically “grown” between the moult stage and the hardening of the new exoskeleton.
Cephalon
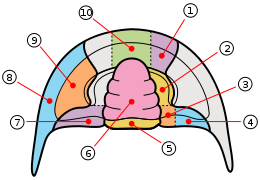
morphological complexity. The glabella forms a dome underneath which sat the “crop” or “stomach”. Generally the exoskeleton has few distinguishing ventral features, but the cephalon often preserves muscle attachment scars and occasionally the hypostome, a small rigid plate comparable to the ventral plate in other arthropods. A toothless mouth and stomach sat upon the hypostome with the mouth facing backwards at the rear edge of the hypostome.
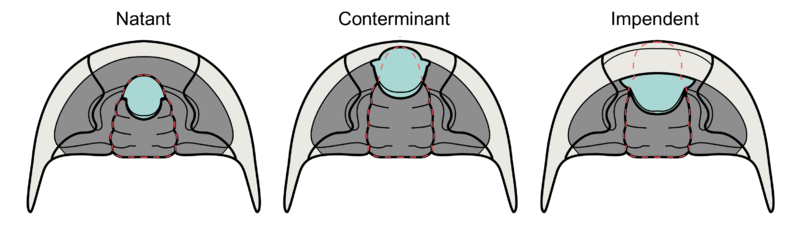
Hypostome morphology is highly variable; sometimes supported by an un-mineralised membrane (natant), sometimes fused onto the anterior doublure with an outline very similar to the glabella above (conterminant) or fused to the anterior doublure with an outline significantly different from the glabella (impendent). Many variations in shape and placement of the hypostome have been described. The size of the glabella and the lateral fringe of the cephalon, together with hypostome variation, have been linked to different lifestyles, diets and specific ecological niches.
The anterior and lateral fringe of the cephalon is greatly enlarged in the Harpetida, in other species a bulge in the pre-glabellar area is preserved that suggests a brood pouch. Highly complex compound eyes are another obvious feature of the cephalon.
Facial sutures
Dorsal sutures
The dorsal surface of the trilobite cephalon (the frontmost tagma, or the ‘head’) can be divided into two regions – the cranidium and the librigena (“free cheeks”). The cranidium can be further divided into the glabella (the central lobe in the cephalon) and the fixigena (“fixed cheeks”). The facial sutures lie along the anterior edge, at the division between the cranidium and the librigena.
Trilobite facial sutures on the dorsal side can be roughly divided into five main types according to where the sutures end relative to the genal angle (the edges where the side and rear margins of the cephalon converge).
- Absent – Facial sutures are lacking in the Olenellina. This is considered a primitive state, and is always combined with the presence of eyes.
- Proparian – The facial suture ends in front of the genal angle, along the lateral margin. Example genera showing this type of suture include Dalmanites of Phacopina (Phacopida) and Ekwipagetia of Eodiscina (Agnostida).
- Gonatoparian – The facial suture ends at the tip of the genal angle. Example genera showing this type of suture include Calymene and Trimerus of Calymenina (Phacopida).
- Opisthoparian – The facial suture ends at the posterior margin of the cephalon. Example genera showing this type of suture include Peltura of Olenina (Ptychopariida) and Bumastus of Illaenina (Corynexochida). This is the most common type of facial suture.
- Hypoparian or marginal – In some trilobites, dorsal sutures may be secondary lost. Several exemplary time series of species show the “migration” of the dorsal suture until it coincides with the margins of the cephalon. As the visual surface of the eye is on the diminishing free cheek (or librigena), the number of lenses tends to go down, and eventually the eye disappears. The loss of dorsal sutures may arise from the proparian state, such as in some Eodiscina like Weymouthia, all Agnostina, and some Phacopina such as Ductina. The marginal sutures exhibited by the harpetids and trinucleioids, however, are derived from opisthoparian sutures. On the other hand, blindness is not always accompanied by the loss of facial sutures.
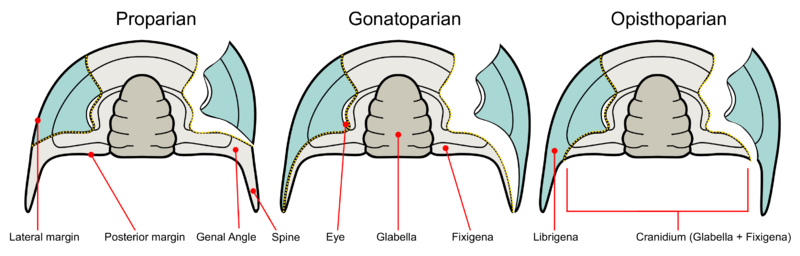
The primitive state of the dorsal sutures is proparian. Opisthoparian sutures have developed several times independently. There are no examples of proparian sutures developing in taxa with opisthoparian ancestry. Trilobites that exhibit opisthoparian sutures as adults commonly have proparian sutures as instars. Hypoparian sutures have also arisen independently in several groups of trilobites.
There are also two types of sutures in the dorsal surface connected to the compound eyes of trilobites.They are:
- Ocular sutures – are sutures surrounding the edges of the compound eye. Trilobites with these sutures lose the entire surface of the eyes when molting. It is common among Cambrian trilobites.
- Palpebral sutures – are sutures which form part of the dorsal facial suture running along the top edges of the compound eye.
Ventral sutures
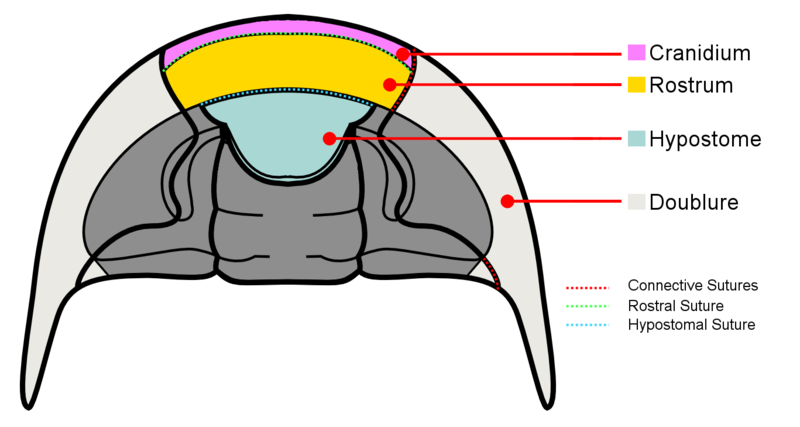
- Connective sutures – are the sutures that continue from the facial sutures past the front margin of the cephalon.
- Rostral suture – is only present when the trilobite possesses a rostrum (or rostral plate). It connects the rostrum to the front part of the dorsal cranidium.
- Hypostomal suture – separates the hypostome from the doublure when the hypostome is of the attached type. It is absent when the hypostome is free-floating (i.e. natant). it is also absent in some coterminant hypostomes where the hypostome is fused to the doublure.
- Median suture – exhibited by asaphid trilobites, they are formed when instead of becoming connective sutures, the two dorsal sutures converge at a point in front of the cephalon then divide straight down the center of the doublure.
Rostrum
The rostrum (or the rostral plate) is a distinct part of the doublure located at the front of the cephalon. It is separated from the rest of the doublure by the rostral suture.
During molting in trilobites like Paradoxides, the rostrum is used to anchor the front part of the trilobite as the cranidium separates from the librigena. The opening created by the arching of the body provides an exit for the molting trilobite.
It is absent in some trilobites like Lachnostoma.
Hypostome
- Natant – Hypostome not attached to doublure. Aligned with front edge of glabella.
- Conterminant – Hypostome attached to rostral plate of doublure. Aligned with front edge of glabella.
- Impendent – Hypostome attached to rostral plate but not aligned to glabella.
Thorax
Each segment consists of the central axial ring and the outer pleurae, which protected the limbs and gills. The pleurae are sometimes abbreviated or extended to form long spines. Apodemes are bulbous projections on the ventral surface of the exoskeleton to which most leg muscles attached, although some leg muscles attached directly to the exoskeleton. Determining a junction between thorax and pygidium can be difficult and many segment counts suffer from this problem.Trilobite fossils are often found “enrolled” (curled up) like modern pill bugs for protection; evidence suggests enrollment helped protect against the inherent weakness of the arthropod cuticle that was exploited by anomalocarid predators.
Some trilobites achieved a fully closed capsule (e.g. Phacops), while others with long pleural spines (e.g. Selenopeltis) left a gap at the sides or those with a small pygidium (e.g. Paradoxides) left a gap between the cephalon and pygidium. In Phacops, the pleurae overlap a smooth bevel (facet) allowing a close seal with the doublure. The doublure carries a Panderian notch or protuberance on each segment to prevent over rotation and achieve a good seal. Even in an agnostid, with only 2 articulating thoracic segments, the process of enrollment required a complex musculature to contract the exoskeleton and return to the flat condition.
Pygidium
The pygidium is formed from a number of segments and the telson fused together. Segments in the pygidium are similar to the thoracic segments (bearing biramous limbs) but are not articulated. Trilobites can be described based on the pygidium being micropygous (pygidium smaller than cephalon), subisopygous (pygidium sub equal to cephalon), isopygous (pygidium equal in size to cephalon), or macropygous (pygidium larger than cephalon).
Prosopon (surface sculpture)
Trilobite exoskeletons show a variety of small-scale structures collectively called prosopon. Prosopon does not include large scale extensions of the cuticle (e.g. hollow pleural spines) but to finer scale features, such as ribbing, domes, pustules, pitting, ridging and perforations. The exact purpose of the prosopon is not resolved but suggestions include structural strengthening, sensory pits or hairs, preventing predator attacks and maintaining aeration while enrolled. In one example, alimentary ridge networks (easily visible in Cambrian trilobites) might have been either digestive or respiratory tubes in the cephalon and other regions.
Spines
Some trilobites such as those of the order Lichida evolved elaborate spiny forms, from the Ordovician until the end of the Devonian period. Examples of these specimens have been found in the Hamar Laghdad Formation of Alnif in Morocco. There is, however, a serious counterfeiting and fakery problem with much of the Moroccan material that is offered commercially. Spectacular spined trilobites have also been found in western Russia; Oklahoma, USA; and Ontario, Canada.
Some trilobites had horns on their heads similar to those of modern beetles. Based on the size, location, and shape of the horns the most likely use of the horns was combat for mates, making the Asaphida family Raphiophoridae the earliest exemplars of this behavior. Another use for these spines was protection from predators. When enrolled, trilobite spines offered additional protection. This conclusion is likely to be applicable to other trilobites as well, such as in the Phacopid trilobite genus Walliserops, that developed spectacular tridents.
Soft body parts
Only 21 or so species are described from which soft body parts are preserved, so some features (e.g. the posterior antenniform cerci preserved only in Olenoides serratus) remain difficult to assess in the wider picture.
Appendages
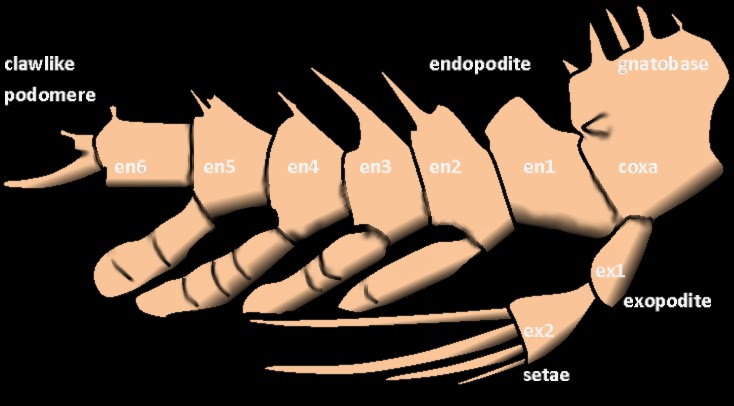
Trilobites had a single pair of preoral antennae and otherwise undifferentiated biramous limbs (2, 3 or 4 cephalic pairs, followed by one pair per thorax segment and some pygidium pairs). Each exopodite (walking leg) had 6 or 7 segments, homologous to other early arthropods. Exopodites are attached to the coxa, which also bore a feather-like endopodite, or gill branch, which was used for respiration and, in some species, swimming. The inside of the coxa (or gnathobase) carries spins, probably to chew prey items. The last exopodite segment usually had claws or spines. Many examples of hairs on the legs suggest adaptations for feeding (as for the gnathobases) or sensory organs to help with walking.
Digestive tract
The toothless mouth of trilobites was situated on the rear edge of the hypostome (facing backwards), in front of the legs attached to the cephalon. The mouth is linked by a small esophagus to the stomach that lay forward of the mouth, below the glabella. The “intestine” led backwards from there to the pygidium. The “feeding limbs” attached to the cephalon are thought to have fed food into the mouth, possibly “slicing” the food on the hypostome and/or gnathobases first. Alternative lifestyles are suggested, with the cephalic legs used to disturb the sediment to make food available. A large glabella, (implying a large stomach), coupled with an impendent hypostome has been used as evidence of more complex food sources, i.e. possibly a carnivorous lifestyle.
Internal organs
While there is direct and implied evidence for the presence and location of the mouth, stomach and digestive tract (see above) the presence of heart, brain and liver are only implied (although “present” in many reconstructions) with little direct geological evidence.
Musculature
Although rarely preserved, long lateral muscles extended from the cephalon to mid way down the pygidium, attaching to the axial rings allowing enrollment while separate muscles on the legs tucked them out of the way.
Sensory organs
Antennae
The pair of antennae suspected in most trilobites (and preserved in a few examples) were highly flexible to allow them to be retracted when the trilobite was enrolled. Also, one species (Olenoides serratus) preserves antennae-like cerci that project from the rear of the trilobite.
Eyes
Trilobite eyes were typically compound, with each lens being an elongated prism. The number of lenses in such an eye varied: some trilobites had only one, while some had thousands of lenses in a single eye. In compound eyes, the lenses were typically arranged hexagonally. The fossil record of trilobite eyes is complete enough that their evolution can be studied through time, which compensates to some extent the lack of preservation of soft internal parts.Lenses of trilobites’ eyes were made of calcite (calcium carbonate, CaCO3). Pure forms of calcite are transparent, and some trilobites used crystallographically oriented, clear calcite crystals to form each lens of each of their eyes. Rigid calcite lenses would have been unable to accommodate to a change of focus like the soft lens in a human eye would; however, in some trilobites the calcite formed an internal doublet structure, giving superb depth of field and minimal spherical aberration, according to optical principles discovered by French scientist René Descartes and Dutch physicist Christiaan Huygens in the 17th century. A living species with similar lenses is the brittle star Ophiocoma wendtii.
In other trilobites, with a Huygens interface apparently missing, a gradient index lens is invoked with the refractive index of the lens changing towards the center.
- Holochroal eyes had a great number (sometimes over 15,000) of small (30–100 μm, rarely larger) lenses. Lenses were hexagonally close packed, touching each other, with a single corneal membrane covering all lenses. Holochroal eyes had no sclera, the white layer covering the eyes of most modern arthropods. Holochroal eyes are the ancestral eye of trilobites, and are by far the most common, found in all orders except the Agnostida, and through the entirety of the Trilobites’ existence. Little is known of the early history of holochroal eyes; Lower and Middle Cambrian trilobites rarely preserve the visual surface. The spatial resolving power of grated eyes (such as holochroal eyes) is dependent on light intensity, circular motion, receptor density, registered light angle, and the extent to which the signal of individual rhabdoms are neurally combined. This implies that lenses need to be larger under low light conditions (such as for Pricyclopyge, when comparing it to Carolinites), and for fast moving predators and prey. As the circular velocity caused by the forward speed of an animal itself is much higher for the ommatidia directed perpendicular to the movement, fast-moving trilobites (such as Carolinites) have eyes flattened from the side and more curved were ommatia are directed to front, back, up and down. Thus eye morphology can be used to make assumptions about the ecosystem of trilobites.
- Schizochroal eyes typically had fewer (around 700), larger lenses than holochroal eyes and are found only in Phacopida. Lenses were separate, with each lens having an individual cornea that extended into a rather large sclera. Schizochroal eyes appear quite suddenly in the early Ordovician, and were presumably derived from a holochroal ancestor. Field of view (all around vision), eye placement and coincidental development of more efficient enrollment mechanisms point to the eye as a more defensive “early warning” system than directly aiding in the hunt for food. Modern eyes that are functionally equivalent to the schizochroal eye were not thought to exist, but are found in the modern insect species Xenos peckii.
- Abathochroal eyes are found only in Cambrian Eodiscina, and have around 70 small separate lenses that had individual cornea. The sclera was separate from the cornea, and did not run as deep as the sclera in schizochroal eyes. Although well preserved examples are sparse in the early fossil record, abathochroal eyes have been recorded in the lower Cambrian, making them among the oldest known. Environmental conditions seem to have resulted in the later loss of visual organs in many Eodiscina.
Secondary blindness is not uncommon, particularly in long lived groups such as the Agnostida and Trinucleioidea. In Proetida and Phacopina from western Europe and particularly Tropidocoryphinae from France (where there is good stratigraphic control), there are well studied trends showing progressive eye reduction between closely related species that eventually leads to blindness.
Several other structures on trilobites have been explained as photo-receptors. Of particular interest are “macula”, the small areas of thinned cuticle on the underside of the hypostome. In some trilobites macula are suggested to function as simple “ventral eyes” that could have detected night and day or allowed a trilobite to navigate while swimming (or turned) upside down.
Sensory pits
There are several types of prosopon that have been suggested as sensory apparatus collecting chemical or vibrational signals. The connection between large pitted fringes on the cephalon of Harpetida and Trinucleoidea with corresponding small or absent eyes makes for an interesting possibility of the fringe as a “compound ear”.
Development

Trilobite larvae are known from the Cambrian to the Carboniferous and from all sub-orders. As instars from closely related taxa are more similar than instars from distantly related taxa, trilobite larvae provide morphological information important in evaluating high-level phylogenetic relationships among trilobites.
Despite the absence of supporting fossil evidence, their similarity to living arthropods has led to the belief that trilobites multiplied sexually and produced eggs. Some species may have kept eggs or larvae in a brood pouch forward of the glabella, particularly when the ecological niche was challenging to larvae. Size and morphology of the first calcified stage are highly variable between (but not within) trilobite taxa, suggesting some trilobites passed through more growth within the egg than others. Early developmental stages prior to calcification of the exoskeleton are a possibility (suggested for fallotaspids), but so is calcification and hatching coinciding.
The earliest post-embryonic trilobite growth stage known with certainty are the “protaspid” stages (anamorphic phase). Starting with an indistinguishable proto-cephalon and proto-pygidium (anaprotaspid) a number of changes occur ending with a transverse furrow separating the proto-cephalon and proto-pygidium (metaprotaspid) that can continue to add segments. Segments are added at the posterior part of the pygidium but, all segments remain fused together.
The “meraspid” stages (anamorphic phase) are marked by the appearance of an articulation between the head and the fused trunk. Prior to the onset of the first meraspid stage the animal had a two-part structure — the head and the plate of fused trunk segments, the pygidium. During the meraspid stages, new segments appeared near the rear of the pygidium as well as additional articulations developing at the front of the pygidium, releasing freely articulating segments into the thorax. Segments are generally added one per moult (although two per moult and one every alternate moult are also recorded), with number of stages equal to the number of thoracic segments. A substantial amount of growth, from less than 25% up to 30%–40%, probably took place in the meraspid stages.
The “holaspid” stages (epimorphic phase) commence when a stable, mature number of segments has been released into the thorax. Moulting continued during the holaspid stages, with no changes in thoracic segment number. Some trilobites are suggested to have continued moulting and growing throughout the life of the individual, albeit at a slower rate on reaching maturity.Some trilobites showed a marked transition in morphology at one particular instar, which has been called “trilobite metamorphosis”. Radical change in morphology is linked to the loss or gain of distinctive features that mark a change in mode of life. A change in lifestyle during development has significance in terms of evolutionary pressure, as the trilobite could pass through several ecological niches on the way to adult development and changes would strongly affect survivorship and dispersal of trilobite taxa. It is worth noting that trilobites with all protaspid stages solely planktonic and later meraspid stages benthic (e.g. asaphids) failed to last through the Ordovician extinctions, while trilobites that were planktonic for only the first protaspid stage before metamorphosing into benthic forms survived (e.g. lichids, phacopids). Pelagic larval life-style proved ill-adapted to the rapid onset of global climatic cooling and loss of tropical shelf habitats during the Ordovician.









{kind=link}