The Carboniferous is a geologic period and system that extends from the end of the Devonian Period, about 358.9 ± 0.4 million years ago, to the beginning of the Permian Period, about 298.9 ± 0.15 Ma. The name Carboniferous means “coal-bearing” and derives from the Latin words carbo (coal) and fero, fers (to carry), and was coined by geologists William Conybeare and William Phillips in 1822. Based on a study of the British rock succession, it was the first of the modern ‘system’ names to be employed, and reflects the fact that many coal beds were formed globally during this time. The Carboniferous is often treated in North America as two geological periods, the earlier Mississippian and the later Pennsylvanian.
Terrestrial life was well established by the Carboniferous period. Amphibians were the dominant land vertebrates, of which one branch would eventually evolve into reptiles, the first fully terrestrial vertebrates. Arthropods were also very common, and many (such as Meganeura), were much larger than those of today. Vast swaths of forest covered the land, which would eventually be laid down and become the coal beds characteristic of the Carboniferous system. A minor marine and terrestrial extinction event occurred in the middle of the period, caused by a change in climate. The later half of the period experienced glaciations, low sea level, and mountain building as the continents collided to form Pangaea.
Subdivisions
In the United States the Carboniferous is usually broken into Mississippian (earlier) and Pennsylvanian (later) Periods. The Mississippian is about twice as long as the Pennsylvanian, but due to the large thickness of coal bearing deposits with Pennsylvanian ages in Europe and North America, the two subperiods were long thought to have been more or less equal. The faunal stages from youngest to oldest, together with some of their subdivisions, are:
Late Pennsylvanian: Gzhelian (most recent)
- Noginskian / Virgilian (part)
- Klazminskian
- Dorogomilovksian / Virgilian (part)
- Chamovnicheskian / Cantabrian / Missourian
- Krevyakinskian / Cantabrian / Missourian
- Myachkovskian / Bolsovian / Desmoinesian
- Podolskian / Desmoinesian
- Kashirskian / Atokan
- Vereiskian / Bolsovian / Atokan
- Melekesskian / Duckmantian
- Cheremshanskian / Langsettian
- Yeadonian
- Marsdenian
- Kinderscoutian
- Alportian
- Chokierian / Chesterian / Elvirian
- Arnsbergian / Elvirian
- Pendleian
- Brigantian / St Genevieve / Gasperian / Chesterian
- Asbian / Meramecian
- Holkerian / Salem
- Arundian / Warsaw / Meramecian
- Chadian / Keokuk / Osagean (part) / Osage (part)
- Ivorian / (part) / Osage (part)
- Hastarian / Kinderhookian / Chouteau
Paleogeography
A mid-Carboniferous drop in sea level precipitated a major marine extinction, one that hit crinoids and ammonites especially hard. This sea level drop and the associated unconformity in North America separate the Mississippian subperiod from the Pennsylvanian subperiod. This happened about 318 million years ago, at the onset of the Permo-Carboniferous Glaciation.
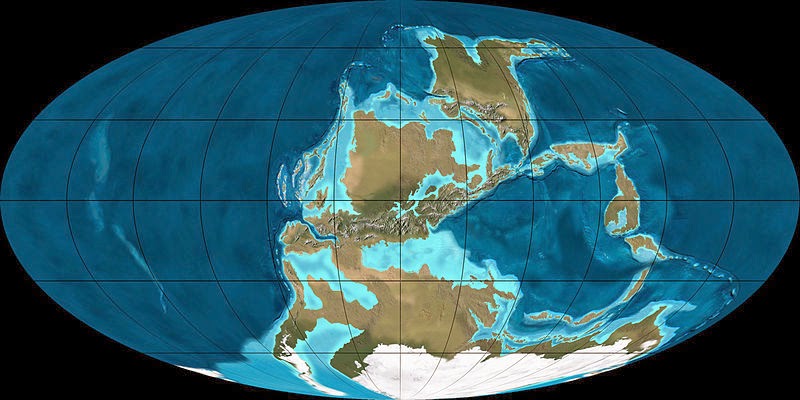
The Carboniferous was a time of active mountain-building, as the supercontinent Pangaea came together. The southern continents remained tied together in the supercontinent Gondwana, which collided with North America–Europe (Laurussia) along the present line of eastern North America. This continental collision resulted in the Hercynian orogeny in Europe, and the Alleghenian orogeny in North America; it also extended the newly uplifted Appalachians southwestward as the Ouachita Mountains. In the same time frame, much of present eastern Eurasian plate welded itself to Europe along the line of the Ural mountains. Most of the Mesozoic supercontinent of Pangea was now assembled, although North China (which would collide in the Latest Carboniferous), and South China continents were still separated from Laurasia. The Late Carboniferous Pangaea was shaped like an “O.”
There were two major oceans in the Carboniferous—Panthalassa and Paleo-Tethys, which was inside the “O” in the Carboniferous Pangaea. Other minor oceans were shrinking and eventually closed – Rheic Ocean (closed by the assembly of South and North America), the small, shallow Ural Ocean (which was closed by the collision of Baltica and Siberia continents, creating the Ural Mountains) and Proto-Tethys Ocean (closed by North China collision with Siberia/Kazakhstania).
Climate
The early part of the Carboniferous was mostly warm; in the later part of the Carboniferous, the climate cooled. Glaciations in Gondwana, triggered by Gondwana’s southward movement, continued into the Permian and because of the lack of clear markers and breaks, the deposits of this glacial period are often referred to as Permo-Carboniferous in age.
The cooling and drying of the climate led to the Carboniferous Rainforest Collapse (CRC). Tropical rainforests fragmented and then were eventually devastated by climate change
Rocks and coal

Carboniferous rocks in Europe and eastern North America largely consist of a repeated sequence of limestone, sandstone, shale and coal beds. In North America, the early Carboniferous is largely marine limestone, which accounts for the division of the Carboniferous into two periods in North American schemes. The Carboniferous coal beds provided much of the fuel for power generation during the Industrial Revolution and are still of great economic importance.
The large coal deposits of the Carboniferous primarily owe their existence to two factors. The first of these is the appearance of bark-bearing trees (and in particular the evolution of the bark fiber lignin). The second is the lower sea levels that occurred during the Carboniferous as compared to the Devonian period. This allowed for the development of extensive lowland swamps and forests in North America and Europe. Based on a genetic analysis of mushroom fungi, David Hibbett and colleagues proposed that large quantities of wood were buried during this period because animals and decomposing bacteria had not yet evolved that could effectively digest the tough lignin. It is assumed that fungi that could break it down did not arise before the end of the period, making future coal formation much more rare. The Carboniferous trees made extensive use of lignin. They had bark to wood ratios of 8 to 1, and even as high as 20 to 1. This compares to modern values less than 1 to 4. This bark, which must have been used as support as well as protection, probably had 38% to 58% lignin. Lignin is insoluble, too large to pass through cell walls, too heterogeneous for specific enzymes, and toxic, so that few organisms other than Basidiomycetes fungi can degrade it. It can not be oxidized in an atmosphere of less than 5% oxygen. It can linger in soil for thousands of years and inhibits decay of other substances. Probably the reason for its high percentages is protection from insect herbivory in a world containing very effective insect herbivores, but nothing remotely as effective as modern insectivores and probably many fewer poisons than currently. In any case coal measures could easily have made thick deposits on well drained soils as well as swamps. The extensive burial of biologically produced carbon led to a buildup of surplus oxygen in the atmosphere; estimates place the peak oxygen content as high as 35%, compared to 21% today. This oxygen level probably increased wildfire activity, as well as resulted in insect and amphibian gigantism—creatures whose size is constrained by respiratory systems that are limited in their ability to diffuse oxygen.
In eastern North America, marine beds are more common in the older part of the period than the later part and are almost entirely absent by the late Carboniferous. More diverse geology existed elsewhere, of course. Marine life is especially rich in crinoids and other echinoderms. Brachiopods were abundant. Trilobites became quite uncommon. On land, large and diverse plant populations existed. Land vertebrates included large amphibians.
Life
Plants
Early Carboniferous land plants, some of which were preserved in coal balls, were very similar to those of the preceding Late Devonian, but new groups also appeared at this time.
The main Early Carboniferous plants were the Equisetales (horse-tails), Sphenophyllales (vine-like plants), Lycopodiales (club mosses), Lepidodendrales (scale trees), Filicales (ferns), Medullosales (informally included in the “seed ferns”, an artificial assemblage of a number of early gymnosperm groups) and the Cordaitales. These continued to dominate throughout the period, but during late Carboniferous, several other groups, Cycadophyta (cycads), the Callistophytales (another group of “seed ferns”), and the Voltziales (related to and sometimes included under the conifers), appeared.
The Carboniferous lycophytes of the order Lepidodendrales, which are cousins (but not ancestors) of the tiny club-moss of today, were huge trees with trunks 30 meters high and up to 1.5 meters in diameter. These included Lepidodendron (with its fruit cone called Lepidostrobus), Halonia, Lepidophloios and Sigillaria. The roots of several of these forms are known as Stigmaria. Unlike present day trees, their secondary growth took place in the cortex, which also provided stability, instead of the xylem. The Cladoxylopsids were large trees, that were ancestors of ferns, first arising in the Carboniferous.
The fronds of some Carboniferous ferns are almost identical with those of living species. Probably many species were epiphytic. Fossil ferns and “seed ferns” include Pecopteris, Cyclopteris, Neuropteris, Alethopteris, and Sphenopteris; Megaphyton and Caulopteris were tree ferns.
The Equisetales included the common giant form Calamites, with a trunk diameter of 30 to 60 cm (24 in) and a height of up to 20 m (66 ft). Sphenophyllum was a slender climbing plant with whorls of leaves, which was probably related both to the calamites and the lycopods.
Cordaites, a tall plant (6 to over 30 meters) with strap-like leaves, was related to the cycads and conifers; the catkin-like inflorescence, which bore yew-like berries, is called Cardiocarpus. These plants were thought to live in swamps and mangroves. True coniferous trees (Walchia, of the order Voltziales) appear later in the Carboniferous, and preferred higher drier ground.
Marine invertebrates

.

In the oceans the most important marine invertebrate groups are the Foraminifera, corals, Bryozoa, Ostracoda, brachiopods, ammonoids, hederelloids, microconchids and echinoderms (especially crinoids). For the first time foraminifera take a prominent part in the marine faunas. The large spindle-shaped genus Fusulina and its relatives were abundant in what is now Russia, China, Japan, North America; other important genera include Valvulina, Endothyra, Archaediscus, and Saccammina (the latter common in Britain and Belgium). Some Carboniferous genera are still extant.
The microscopic shells of radiolarians are found in cherts of this age in the Culm of Devon and Cornwall, and in Russia, Germany and elsewhere. Sponges are known from spicules and anchor ropes, and include various forms such as the Calcispongea Cotyliscus and Girtycoelia, the demosponge Chaetetes, and the genus of unusual colonial glass sponges Titusvillia.
Both reef-building and solitary corals diversify and flourish; these include both rugose (for example, Caninia, Corwenia, Neozaphrentis), heterocorals, and tabulate (for example, Chladochonus, Michelinia) forms. Conularids were well represented by Conularia
Bryozoa are abundant in some regions; the fenestellids including Fenestella, Polypora, and Archimedes, so named because it is in the shape of an Archimedean screw. Brachiopods are also abundant; they include productids, some of which (for example, Gigantoproductus) reached very large (for brachiopods) size and had very thick shells, while others like Chonetes were more conservative in form. Athyridids, spiriferids, rhynchonellids, and terebratulids are also very common. Inarticulate forms include Discina and Crania. Some species and genera had a very wide distribution with only minor variations.
Annelids such as Serpulites are common fossils in some horizons. Among the mollusca, the bivalves continue to increase in numbers and importance. Typical genera include Aviculopecten, Posidonomya, Nucula, Carbonicola, Edmondia, and Modiola Gastropods are also numerous, including the genera Murchisonia, Euomphalus, Naticopsis. Nautiloid cephalopods are represented by tightly coiled nautilids, with straight-shelled and curved-shelled forms becoming increasingly rare. Goniatite ammonoids are common.
Trilobites are rarer than in previous periods, on a steady trend towards extinction, represented only by the proetid group. Ostracoda, a class of crustaceans, were abundant as representatives of the meiobenthos; genera included Amphissites, Bairdia, Beyrichiopsis, Cavellina, Coryellina, Cribroconcha, Hollinella, Kirkbya, Knoxiella, and Libumella.
Amongst the echinoderms, the crinoids were the most numerous. Dense submarine thickets of long-stemmed crinoids appear to have flourished in shallow seas, and their remains were consolidated into thick beds of rock. Prominent genera include Cyathocrinus, Woodocrinus, and Actinocrinus. Echinoids such as Archaeocidaris and Palaeechinus were also present. The blastoids, which included the Pentreinitidae and Codasteridae and superficially resembled crinoids in the possession of long stalks attached to the seabed, attain their maximum development at this time.
Freshwater and lagoonal invertebrates
Freshwater Carboniferous invertebrates include various bivalve molluscs that lived in brackish or fresh water, such as Anthraconaia, Naiadites, and Carbonicola; diverse crustaceans such as Candona, Carbonita, Darwinula, Estheria, Acanthocaris, Dithyrocaris, and Anthrapalaemon.
The Eurypterids were also diverse, and are represented by such genera as Eurypterus, Glyptoscorpius, Anthraconectes, Megarachne (originally misinterpreted as a giant spider) and the specialised very large Hibbertopterus. Many of these were amphibious.
Frequently a temporary return of marine conditions resulted in marine or brackish water genera such as Lingula, Orbiculoidea, and Productus being found in the thin beds known as marine bands.
Terrestrial invertebrates
Fish
Many fish inhabited the Carboniferous seas; predominantly Elasmobranchs (sharks and their relatives). These included some, like Psammodus, with crushing pavement-like teeth adapted for grinding the shells of brachiopods, crustaceans, and other marine organisms. Other sharks had piercing teeth, such as the Symmoriida; some, the petalodonts, had peculiar cycloid cutting teeth. Most of the sharks were marine, but the Xenacanthida invaded fresh waters of the coal swamps. Among the bony fish, the Palaeonisciformes found in coastal waters also appear to have migrated to rivers. Sarcopterygian fish were also prominent, and one group, the Rhizodonts, reached very large size.
Most species of Carboniferous marine fish have been described largely from teeth, fin spines and dermal ossicles, with smaller freshwater fish preserved whole.
Freshwater fish were abundant, and include the genera Ctenodus, Uronemus, Acanthodes, Cheirodus, and Gyracanthus.
Sharks (especially the Stethacanthids) underwent a major evolutionary radiation during the Carboniferous. It is believed that this evolutionary radiation occurred because the decline of the placoderms at the end of the Devonian period caused many environmental niches to become unoccupied and allowed new organisms to evolve and fill these niches. As a result of the evolutionary radiation carboniferous sharks assumed a wide variety of bizarre shapes including Stethacanthus which possessed a flat brush-like dorsal fin with a patch of denticles on its top. Stethacanthus’ unusual fin may have been used in mating rituals.
Tetrapods
Carboniferous amphibians were diverse and common by the middle of the period, more so than they are today; some were as long as 6 meters, and those fully terrestrial as adults had scaly skin. They included a number of basal tetrapod groups classified in early books under the Labyrinthodontia. These had long bodies, a head covered with bony plates and generally weak or undeveloped limbs. The largest were over 2 meters long. They were accompanied by an assemblage of smaller amphibians included under the Lepospondyli, often only about 15 cm (6 in) long. Some Carboniferous amphibians were aquatic and lived in rivers (Loxomma, Eogyrinus, Proterogyrinus); others may have been semi-aquatic (Ophiderpeton, Amphibamus, Hyloplesion) or terrestrial (Dendrerpeton, Tuditanus, Anthracosaurus).
The Carboniferous Rainforest Collapse slowed the evolution of amphibians who could not survive as well in the cooler, drier conditions. Reptiles, however prospered due to specific key adaptations. One of the greatest evolutionary innovations of the Carboniferous was the amniote egg, which allowed for the further exploitation of the land by certain tetrapods. These included the earliest sauropsid reptiles (Hylonomus), and the earliest known synapsid (Archaeothyris). These small lizard-like animals quickly gave rise to many descendants. The amniote egg allowed these ancestors of all later birds, mammals, and reptiles to reproduce on land by preventing the desiccation, or drying-out, of the embryo inside.
Reptiles underwent a major evolutionary radiation in response to the drier climate that proceeded the rainforest collapse. By the end of the Carboniferous period, amniotes had already diversified into a number of groups, including protorothyridids, captorhinids, aeroscelids, and several families of pelycosaurs.
Fungi
Extinction events
Romer’s gap
Carboniferous rainforest collapse
Before the end of the Carboniferous Period, an extinction event occurred. On land this event is referred to as the Carboniferous Rainforest Collapse (CRC). Vast tropical rainforests collapsed suddenly as the climate changed from hot and humid to cool and arid. This was likely caused by intense glaciation and a drop in sea levels.
The new climatic conditions were not favorable to the growth of rainforest and the animals within them. Rainforests shrank into isolated islands, surrounded by seasonally dry habitats. Towering lycopsid forests with a heterogeneous mixture of vegetation were replaced by much less diverse tree-fern dominated flora.
Amphibians, the dominant vertebrates at the time, fared poorly through this event with large losses in biodiversity; reptiles continued to diversify due to key adaptations that let them survive in the drier habitat, specifically the hard-shelled egg and scales both of which retain water better than their amphibian counterparts.









{kind=link}