Tetrapod is the name given to any vertebrate animal with four (tetra) legs (pod). There are more than 30,000 living species of tetrapod known today, and this includes many of the animals we are familiar with like mammals, crocodiles, snakes, and birds. The question of how they reached this level of diversity is still very much open, and we know that by peering back into pre-history with the fossil record the number of tetrapod species has not remained constant through time.
The history of tetrapod life is punctuated by episodes of mass extinction, huge reorganisations of life both in terms of ecology and the relative dominance of different groups. With extinction comes radiation, the origin of new groups to fill the vacant environments and ecologies left as species die out. This natural variation in the fluctuating number of species through time is inconsistent through time, and unlocking what drives such patterns remains one of the long standing, but well-investigated, realms of palaeontological research.
The importance of why we research such things might not be immediately clear, but there are two main reasons that come to mind:
Understanding the evolution of life on this planet is a totally awesome thing to study;
By learning about past patterns and processes in the history of life, we might better understand how to stop destroying it so much in the future.
Recently, we have been able to provide some answers to the questions of how diverse through time has life been, based on the building of large fossil occurrence databases and new methods of analysing them. One such development has been the Paleobiology Database, a professional crowd-sourced archive of fossil history, where the context of fossils is provided in both space and time, and largely based on the published record of fossil discoveries.
Unfortunately, simply counting the number of species preserved through time as fossils is not a good way of measuring biodiversity: we can’t simply say we have n species found in this time period, and therefore diversity was n. The reasons for this are due to differences in how the fossil record is preserved through time, and presented to us as collectors and researchers. For example, what do you do if you have a lot of fossiliferous rock in one time period, but very little in another, and does an absence of fossil-bearing rock imply low diversity or something else? Or how do you compare samples if you have 100 fossil occurrences representing 20 species in one time bin, but 150 occurrences representing 50 species in another? This is the fundamental problem with the evenness of sampling, which is sometimes called ‘sampling bias’, and has plagued palaeontologists since the day someone was kind enough to point it out.
However, this problem, as with all of those in science, is not completely intractable! Thankfully, there exist a whole suite of methods to counter this issue of sampling variation. One of the more recently implemented of these is one called Shareholder Quorum Subsampling (SQS) developed by John Alroy, the statistical sensei of palaeontology. It has a horrific name, yes, but SQS is essentially a method that weights how diverse one species is (the shareholder) based on its frequency of occurrence. Using this, and a calculation taken from ecology of how evenly sampled your overall sample is, you can then count through repeated random trials (subsampling) until a pre-defined threshold (the quorum; you see where this is going now..) applied to all samples is met. If that doesn’t make sense, don’t worry about it – it took me years to understand and implement this method, and that was only with the help of John himself (who, incidentally, is an author on the paper this post is all about!)
By applying SQS with our development of large fossil occurrence datasets, voila, we are able to gain renewed insight into the diversity of life through history in a way that accounts for the inherent biases of the fossil record!
And that’s just what a new study in PLOS Biology set out to do. Led by Roger Benson of the University of Oxford, an international team of researchers applied SQS to one of the largest tetrapod fossil occurrence databases ever assembled (if not the largest!), comprising more than 27,000 individual fossil occurrences! This represented almost 5000 fossil species, and the data were restricted to just those fossils that dwelled on land – so this excludes groups like ichthyosaurs and plesiosaurs, for example. They also excluded flying tetrapods, so birds, bats and mammals, as these are known to have very different preservational histories in the fossil record. For palaeontology though, this is definitely ‘big data’.
The team restricted their analyses to just the Mesozoic to early Paleogene, a time span of around 190 million years (a fairly long time, even by geological standards!). If you think about it, that’s 5000 species over about 190 million years, which compared to 30,000 around today is pretty weird even in itself.
But anyway, armed with this hefty dataset, it becomes possible to group them by the time at which they occur in the fossil record, and use SQS to assess how their diversity changes through time. Simples?
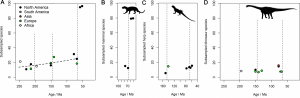
What did they find? Well, perhaps a quite unexpected result. They found that tetrapod diversity actually remained fairly constant and stable throughout most of this time period. This was in spite of the fact that we have new groups originating and diversifying, such as mammals, amphibians, and dinosaurs, while other groups go extinct or massively reduce in numbers. In all, species diversity of non-flying mammals increased less than double, which suggests that there is some suppressing or limiting factor that controls long term diversification in tetrapods.
Interestingly, the extent of sampling bias detected varied substantially on a regional level, based on approximate palaeocontinental reconstructions (the world was a very different place 100 million years ago!) What this means is that global diversity counts, when applying techniques like SQS, are poor reflectors of what is happening to diversity at a regional level through time.
The team also found an anomaly at the end-Cretaceous mass extinction, with a four-fold expansion of species diversity following the extinction, suggesting that this extinction was highly important in paving the way for the origins of modern biodiversity. This result does not seem to be due to the aforementioned geographical biases, and instead appears to be almost entirely due to an explosive radiation of mammals subsequent to the extinction event. This led to an almost total restructuring of pre-extinction terrestrial ecosystems, and ultimately led to what is commonly referred to as ‘the rise of the mammals’. Sorry dinosaurs!
However, Benson and colleagues to advise caution in interpreting this as a genuine evolutionary phenomenon. This is because mammalian species are much more easily identifiable based on the remains of teeth, and therefore part of this apparent increase in diversity after the extinction event might simply be due to changes in how vertebrate discovery and taxonomy works.
Overall, the results obtained in the study show that tetrapod diversity throughout the Mesozoic less than doubled when sampling issues are accounted for. This in turn implies a very low, near-zero rate of diversification through time. Such a discovery is highly at odds with previous analyses which either accounted for sampling issues in different ways or ignored them entirely, which suggested that tetrapod diversity actually was unconstrained and rapidly increased on land throughout the Mesozoic.
The reasons for why this pattern exists though are more complicated. It is difficult to tell whether it is due to the achievement of a diversity equilibrium, where extinction rates and rates of diversification balance each other out, a phenomenon often referred to as ‘diversity dependence’, or perhaps simply due to overall stability in environments throughout the Mesozoic. The result of both of these hypotheses would be static diversity through time. Alternatively, it could be something a little more exciting, where we have a scenario in which diversifications are largely unconstrained, but regulated by more frequent episodes of extinction or dramatic reductions in diversification rates. So, in short, it’s a bit of a mess and quite difficult to tell.
This is already a rather long post, and could go on forever about the cool results obtained in the Benson et al. study. So I’ll end with a couple of key points:
The fossil record is our gateway to understanding the patterns and processes regulating past diversity, and remains key to understanding the origins and future of modern biodiversity;
Mesozoic tetrapod faunas appear to have been relatively stable when regional sampling biases are accounted for, and document very low diversification rates;
Palaeontology has never been more relevant in helping us to understand the evolution of life, and we’re making huge leaps forward in terms of data and methods to realise this.
Reference:
Roger B. J. Benson et al. Near-Stasis in the Long-Term Diversification of Mesozoic Tetrapods, PLOS Biology (2016). DOI: 10.1371/journal.pbio.1002359
Note: The above post is reprinted from materials provided by PLOS Blogs.









 legs (pod). There are more than 30,000 living species of tetrapod known today, and thi){kind=link}